 فارسی
فارسی
 English
English


انواع استرس محیطی در پرورش طیور
چکیده
استرسهای غیرزیستی (abiotic stressors) در صنعت پرورش طیور، از جمله نوسانات دمایی، صدا، ارتعاشات و مدیریت نادرست نور، تأثیرات منفی بر رفاه، رشد و بهرهوری پرندگان میگذارند. این مقاله، با ارجاع به مقالات بروز جهانی بر اساس مرورهای اخیر مانند Ncho et al., ۲۰۲۴، مکانیسمهای فیزیولوژیک، اثرات و تعاملات این عوامل را بررسی میکند. همچنین، نگاهی بر تعاملات عوامل استرسزا و استراتژیهای مدیریتی صورت گرفته است.
با توجه به تغییرات اقلیمی، پیشبینی میشود نوسانات حرارتی تا ۲۰۵۰ تا ۳۰% افزایش یابد، که نیاز به رویکردهای پایدار را برجسته میسازد.
مرور مبانی نظری استرس محیطی
در دامپروری مدرن، استرس را میتوان بهعنوان یک پاسخ خودکار ناشی از قرار گرفتن در معرض شرایط نامساعد محیطی در نظر گرفت. این پاسخ میتواند از ناراحتی خفیف تا پیامدهای شدید، از جمله مرگومیر، متغیر باشد. صنعت طیور که نقش چشمگیری در تغذیه انسان دارد، از این مشکل مستثنا نیست. گرچه انتخاب ژنتیکی طی چند دهه برای افزایش بهرهوری تولید بهکار گرفته شده است، اما این فرایند موجب کاهش مقاومت نسبت به استرس نیز شده است. استرس از راه مجموعهای از واکنشهای فیزیولوژیک بروز مییابد؛ از جمله شناسایی عامل استرسزا، فعال شدن دستگاه عصبی سمپاتیک و غده آدرنال مدولا و در ادامه ترشح انواع هورمونها متعاقب آن.
هرچند دورههای کوتاه استرس قابل تحملاند، اما قرارگیری طولانیمدت در معرض استرس، میتواند پیامدهای وخیمتری داشته باشد. برای مثال، نوسان شدید دمای محیط میتواند موجب تجمع گونههای فعال اکسیژن (ROS)، اختلال در عملکرد تولیدمثلی و کاهش ایمنی گردد. همچنین، لرزشهای مکانیکی تأثیر منفی بر کیفیت گوشت جوجههای گوشتی در زمان حملونقل و نیز بر کیفیت و جوجهدرآوری تخمها در جوجهکشیها دارد. تولید تخممرغ بهشدت تحت تأثیر شدت و الگوی نور است و مدیریت نادرست نور میتواند منجر به کمبودهایی همچون ناهنجاریهای بینایی، تغییر شکلهای اسکلتی و مشکلات گردش خون شود. اگرچه شواهد فزایندهای وجود دارد که نشاندهنده اثر عوامل استرسزای محیطی بر فیزیولوژی طیور است، اما توزیع این عوامل در پژوهشها نامتوازن است.
استرس در حیوانات را میتوان بهعنوان پاسخ فیزیولوژیکی و روانی به عوامل داخلی و خارجی تعریف کرد که تعادل هموستاتیکی یا فیزیولوژیکی طبیعی حیوان را بر هم میزنند. بصورت کلی این عوامل میتوانند شامل مشکلات محیط زیستی، تغییرات دما، در دسترس بودن غذا، تعاملات اجتماعی و قرار گرفتن در معرض شکارچیان یا سایر تهدیدها باشند. در پاسخ به استرس، حیوانات معمولاً مجموعهای از تغییرات فیزیولوژیک را تجربه میکنند، از جمله ترشح هورمونهای استرس مانند کورتیزول در پستانداران یا کورتیکوسترون در پرندگان. این هورمونها آثار گستردهای بر بدن حیوان دارند و هدف از ترشح آنها، کمک به حیوان برای سازگاری با چالشهای فوری است. با این حال، استرس مزمن یا بیش از حد میتواند آثار زیانبار بر سلامت و تندرستی حیوان داشته باشد.
علائم شایع استرس در حیوانات شامل تغییر رفتار، تغییر در اشتها، کاهش وزن، کاهش موفقیت تولیدمثل، تضعیف عملکرد ایمنی و افزایش حساسیت به بیماریها است و در زمینه پرورش طیور، مجموعهای از عوامل گوناگون موجب شکلگیری شبکهای پیچیده از استرسها میشوند. برای مثال، عوامل استرسزای محیطی مانند نوسانات دمایی، میزان رطوبت، کیفیت هوا و شرایط نوری میتوانند سلامت، رشد و بازدهی طیور را بهشدت تحت تأثیر قرار دهند و در مقابل تنظیم این عوامل متناسب با نیازهای سویه میتواند تأثیرات مثبتی بر سلامت و عملکرد آنها بگذارد.
برخی از مطالعات برای کاهش اثرات عوامل استرسزا
- بر اساس گزارش گروهی از محققین، استرس گرمایی اثر نامطلوبی بر رفاه و بهرهوری طیور دارد و دستکاری حرارتی در طی دوره جوجهکشی و قرار دادن تخمها در دمای بالاتر (۳۹.۵ درجه سانتیگراد با ۶۰% رطوبت نسبی) از روز ۱۲ تا ۱۸ سبب بهبود رشد جنینی، ویژگیهای هچ و کیفیت جوجهها میشود. پس از خروج از تخم، این جوجهها تحمل حرارتی بهتر و وزن بدن بالاتری نشان میدهند. این گروه توصیه میکنند دستکاری حرارتی در روزهای ۱۲ تا ۱۸ جوجهکشی برای بهرهمندی از اثرات مثبت آن بر عملکرد پیش و پس از هچ و نیز بهبود بهرهوری و رفاه طیور انجام شود.
- در مطالعهای مشابه با بهکارگیری نور LED بهجای لامپهای رشتهای برای مرغهای مادر گوشتی، بهبود چشمگیری در بهرهوری شامل وزن بدن، ضریب تبدیل خوراک و تولید تخممرغ گزارش کردهاند. این مزایا در طول دوره تولید نیز تداوم داشت و نور LED را به گزینهای بسیار مؤثر برای ارتقای بهرهوری و رفاه طیور تبدیل کرد.
عوامل استرسزای مدیریتی با شیوههای مدیریتی فارم در ارتباط مستقیم بوده و جنبههایی همچون طراحی سالن، روشهای جابجایی و راهبردهای تغذیه را دربر میگیرند. ازدحام بیش از حد یا تهویه ناکافی در سالنها میتواند محیطی استرسزا ایجاد کرده و زمینهساز بروز بیماری شود. جابجایی نامناسب در حین حملونقل یا در فرآیندهای روزمره نیز میتواند اثرات منفی بر رفاه پرندگان داشته باشد. همچنین تغذیه موردی حیاتی است، زیرا جیرههای نامتعادل میتوانند موجب رشد ناکافی و کاهش بازده تولید شوند. مدیریت مؤثر این استرسها برای تضمین سلامت کلی طیور و بهینهسازی نتایج تولید، امری حیاتی است. برای اتخاذ تصمیمات آگاهانه و تدوین راهبردهای مؤثر در زمینه مدیریت و کاهش عوامل بالقوه استرسزا، درک پیچیدگی این عوامل ضروری است.
هماهنگی پاسخ استرس در حیوانات
ادغام و پردازش استرس در حیوانات، فرآیندی پیچیده و بسیار هماهنگ است که شامل تعامل سیستمهای فیزیولوژیک و عصبی گوناگون برای پاسخ به تهدیدها یا چالشهای ادراکشده میباشد. این پاسخ استرسی شامل واکنش بدن در برابر عوامل استرسزای درونی و بیرونی بوده و به مجموعهای از رویدادهای درهمتنیده در بدن حیوان منجر میشود. واکنش به عوامل استرسزا با شناسایی و ارسال سیگنالِ تهدیدِ ادراکشده در سازوکارهای زیستی حیوان آغاز میشود.
در ادامه، این فرآیند موجب فعال شدن سازوکارهای نوروفیزیولوژیکی میگردد که هدف آن آغاز پاسخ زیستی برای مقابله و کاهش آسیبهای احتمالی است. شایان توجه است که گیرندههای حسی گوناگون نهتنها اطلاعاتی درباره عامل استرسزا دریافت میکنند، بلکه این دادهها را به سیگنالهای عصبی تبدیل مینمایند. این سیگنالها سپس به مراکز شناختی و غیرشناختی دستگاه عصبی منتقل میشوند و زمینهساز ایجاد واکنشی هماهنگ در برابر چالش پیشآمده میگردند. این پاسخ هماهنگ ناشی از تعامل میان دستگاه عصبی مرکزی (CNS)، دستگاه درونریز (غدد درونریز) و دستگاه ایمنی است که در مجموع به محرکهای استرسزا واکنش نشان داده و رفتار حیوان را تحت تأثیر قرار میدهند (مطابق شکل ۱).

فعال شدن محور HPA در مغز
وجود هورمونها، انتقالدهندههای عصبی و گیرندههای مشترک میان این سه سامانه، بیانگر وجود ارتباط و تعامل میان آنهاست. دریافت اولیه یک عامل استرسزا، پاسخ پیچیدهای را در حیوانات برمیانگیزد که مبتنی بر سیستمهای حسی برای شناسایی تغییرات محیطی یا تهدیدهای بالقوه است. اطلاعات حسی سپس به نواحی خاصی از مغز مانند آمیگدال و قشر پیشپیشانی ارسال میشود که اهمیت و خطر بالقوه عامل استرسزا را ارزیابی میکنند. این نواحی مغزی نقش اساسی در تعیین نیاز یا عدم نیاز به پاسخ استرس ایفا میکنند.
بهمحض اینکه مغز عامل استرسزا را مهم تشخیص دهد، محور هیپوتالاموس-هیپوفیز-آدرنال (HPA) که یک سامانه تنظیمی بنیادین در بدن است، فعال میشود. هیپوتالاموس که ساختاری کوچک اما حیاتی در مغز است، هورمون آزادکننده کورتیکوتروپین ترشح میکند که از طریق سامانه پورتال هیپوفیزی به بخش قدامی هیپوفیز میرسد. در هیپوفیز قدامی، این هورمون ترشح و سنتز هورمونهایی چون آدرنوکورتیکوتروپیک (ACTH)، بتا‑اندورفین، بتا‑لیپوتروپین و آلفا‑هورمون محرک ملانوسیت (α‑MSH /α‑melanotropin) را تحریک میکند. افزایش سطح گلوکوکورتیکوئیدها در خون میتواند بهصورت بالقوه ترشح ACTH از غده هیپوفیز را مهار کند؛ این فرآیند نمونهای از سازوکار کنترل بازخوردی محسوب میشود.
با این حال، ترشح ACTH وابسته به شدت عامل استرسزا بوده و تحت تأثیر سیستم بازخوردی حساس و تنظیمشدهای قرار دارد. در نتیجه، استرسهای خفیف ممکن است بهتدریج از طریق بازخورد گلوکوکورتیکوئیدی مهار شوند، در حالیکه استرسهای شدیدتر کمتر قابل سرکوب هستند. ترشح هورمونهای استرس مانند کورتیکوسترون اثرات عمیق و گستردهای را در سراسر بدن ایجاد میکند. این تغییرات فیزیولوژیکی بهمنظور آمادهسازی حیوان جهت پاسخ فوری به تهدید ادراکشده انجام میگیرد.
تغییرات فیزیولوژیک برای پاسخ به استرس
افزایش سطح گلوکز خون منبع سریع انرژی برای پاسخ «مبارزه یا گریز» فراهم میکند. افزونبراین، ضربان قلب و فشار خون افزایشیافته و انتقال اکسیژن کافی به بافتهای حیاتی تضمین میشود. دستگاه تنفسی با اتساع برونشیولها واکنش نشان میدهد تا جذب اکسیژن افزایش یابد. با این حال، دستگاه گوارش دستخوش تغییرات موقتی میشود، بهگونهایکه هضم و جذب مواد غذایی متوقف شده و منابع به قسمتهای دیگر بدن هدایت میگردند.
استرس همچنین میتواند بهطور موقت پاسخ دستگاه ایمنی را سرکوب کرده و حیوان را نسبت به عفونتها مستعدتر سازد. نشان داده شده است که عوامل استرسزای فیزیکی و روانی فعالیت لنفوسیتهای T و B و همچنین سلولهای کشنده طبیعی را مهار میکنند. افزونبراین، این عوامل میتوانند ساخت برخی سایتوکاینها نظیر (اینترلوکین‑۲ IL‑۲) و (اینترفرون‑گاما IFN‑γ) را کاهش دهند. سایتوکاینهایی همچون IL‑۱، IL‑۶، فاکتور نکروز تومور آلفا TNF‑α و IFN‑γ نقش حیاتی بهعنوان میانجی در پاسخهای ایمنی و پاتولوژیک ناشی از استرس و عفونت دارند.
با این حال، هیپوکامپ که ناحیهای حیاتی در مغز است، نقش تعدیلکنندهای ایفا کرده و با ارائه بازخورد منفی به محور HPA، به پایان یافتن پاسخ استرس پس از رفع تهدید کمک میکند. مواجهه مزمن یا تکرارشونده با عوامل استرسزا میتواند آثار درازمدتی بر نوروبیولوژی حیوان داشته باشد و موجب تغییرات ساختاری در مغز بهویژه در نواحی چون آمیگدال و هیپوکامپ شود. پژوهشهای اخیر (Wang et al., ۲۰۲۵) نشان میدهند که این تغییرات با اختلال میکروبیوم روده مرتبط است، که تعادل HPA را بیشتر برهم میزند.
عوامل استرسزای مرتبط با دما
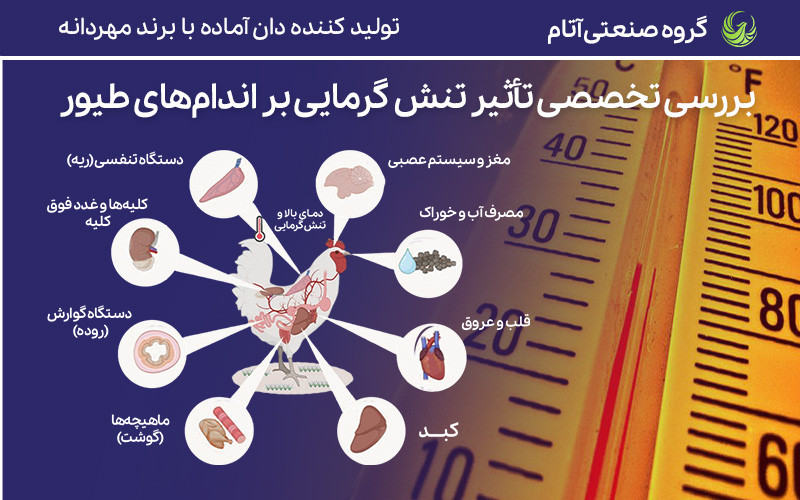
در زمینه پرورش طیور، دما در سراسر مراحل رشد آنها از مرحله جنینی تا بلوغ نقشی محوری ایفا میکند. گونههای معمول طیور توان محدودی در تنظیم دمای بدن خود دارند. این ویژگی باعث میشود طیور، بهویژه نژادهای مدرنِ حاصل از سالها اصلاح نژادی انتخابی، نسبت به دماهای محیطی بالا و پایین حساس و آسیبپذیر باشند. برای نمونه در طول رشد جنینی، بسیاری از گونههای مورد استفاده در تولید دام و طیور به دماهایی بالاتر از ۳۶ درجه سانتیگراد نیاز دارند. افزون بر این، انحراف از دمای بهینه بهویژه قرار گرفتن در معرض دمای پایین در هفته اول پس از هچ، میتواند منجر به بروز مشکلات شدید سلامتی و در نهایت مرگومیر شود. در مقابل برای مثال، جوجههای گوشتی پس از ۲۷ روزگی در بازه ۲۰ تا ۲۴ درجه سانتیگراد بیشترین رشد مطلوب را نشان میدهند. بنابراین، درک اثرات فیزیولوژیک نوسانات شدید دما بر بدن پرندگان امری بنیادین و حیاتی است.
هر دو نوع استرس گرمایی و سرمایی آثار زیانباری دارند (شکل ۲) و این امر بر ضرورت شناختِ رابطه پیچیده میان نوسانات دما و فیزیولوژی پرندگان تأکید میکند. با توجه به تغییرات اقلیمی، مدلهای IPCC (۲۰۲۵) پیشبینی میکنند افزایش ۱.۵ درجهای دما تا ۲۰۵۰، که استرس گرمایی را در مناطق گرمسیری ۲۵% تشدید کند.

1) استرس گرمایی در طیور
استرس گرمایی زمانی رخ میدهد که سازوکارهای طبیعی خنکسازی بدن نتوانند حرارت را بهطور مؤثر دفع کنند و در نتیجه دمای بدن بهصورت محسوس افزایش مییابد. برخلاف پستانداران، پرندگان فاقد توانایی تعریق هستند؛ ازاینرو ظرفیت آنها در خنک شدن در محیطهای گرم بهشدت محدود است. در زمان بروز استرس گرمایی، تعامل پیچیدهای میان تغییرات رفتاری، ایمنی و فیزیولوژیک در بدن پرنده شکل میگیرد که بهطور عمیق بر سلامت و عملکرد تولیدی گونههای پرندگان تأثیر میگذارد. این تغییرات در واکنش به افزایش دما چندبعدی بوده و شامل مجموعهای از دگرگونیهای مرتبط با متابولیسم هستند. اخیراً، مطالعات نشان دادهاند که استرس گرمایی بر میکروبیوم روده تأثیر گذاشته و تنوع باکتریهای مفید را کاهش میدهد، که این امر ایمنی و هضم را مختل میکند.
پاسخهای رفتاری طیور به دمای بالای محیطی
یکی از مهمترین تغییرات رفتاری مشاهدهشده در پرندگانِ تحت استرس گرمایی، تغییر در میزان فعالیت حرکتی آنهاست. برای کاهش تولید گرما، پرندگان تمایل دارند حرکات خود را محدود کرده و ترجیح میدهند در وضعیت ثابت باقی بمانند. یافتههای قابلتوجه از مطالعهای که با پایش تصویری ۲۴ساعته انجام شد، نشان داد جوجههای گوشتی در معرض دماهای بالای محیطی طولانیمدت، عمدتاً حالت نشسته و بیتحرک اتخاذ کرده و تنها در مواقع ضروری، عمدتاً برای دسترسی به منابع آب از جای خود بلند میشوند. علاوه بر این، عمدتاً بهدلیل گرمای تولیدشده در جریان فرآیند خوردن غذا، میزان مصرف خوراک بهطور محسوسی کاهش مییابد. این کاهش مصرف خوراک، نهتنها رشد پرنده را مختل میکند، بلکه جذب مواد مغذی ضروری را بهشدت محدود کرده و در نتیجه رشد کلی و بهرهوری تولید را تحت تأثیر قرار میدهد.
برعکس، برخی رفتارهای خاص در دورههای استرس گرمایی تشدید میشوند. برای نمونه، مصرف آب در پرندگانِ تحت استرس گرمایی افزایش مییابد. آزمایشهای اخیر نشان دادهاند که جوجههای گوشتی در معرض شرایط استرس گرمایی تناوبی، نسبت آب به خوراک خود را بیش از ۳۰% افزایش میدهند؛ این امر نقش حیاتی آبرسانی در کاهش اثرات استرس گرمایی را برجسته میسازد. افزایش مصرف آب برای حفظ تعادل بهینه آب بدن حیاتی است و به برقراری عملکردهای حیاتی فیزیولوژیک در شرایط حرارتی دشوار کمک میکند.
همچنین مشاهدات نشان میدهد که پرندگان برای بهبود خنکسازی بدن، حالات بدنی ویژهای به خود میگیرند. گشودن بالها و تنفس دهانی (لهله زدن) بیش از حد، از رفتارهای رایج در هنگام افزایش دما به بیش از حد مطلوب بهشمار میروند. در فارمهای بدون قفس، پرندگان بهطور غریزی خود را با بستر پوشانده و لایهای ایجاد میکنند که در خنکسازی تبخیری مؤثر است. این سازوکارهای رفتاری ظریف، نقشی کلیدی در توان سازگاری پرندگان با استرس گرمایی ایفا کرده و شیوههای خاص واکنش آنها به چالشهای محیطی را آشکار میسازد.
پاسخهای فیزیولوژیک طیور به دمای بالای محیط
دومین دسته از تغییراتی که با استرس گرمایی مرتبط هستند، تغییرات فیزیولوژیکی میباشند. تغییرات فیزیولوژیکی در بدن پرندگان میتوانند بهطور چشمگیری عملکردها و فرایندهای مختلف بدن را مختل سازند. یکی از جنبههای حیاتی این تغییرات فیزیولوژیکی، القای استرس اکسیداتیو ناشی از گرما است. استرس اکسیداتیو از عدم تعادل میان رادیکالهای آزاد و سیستم آنتیاکسیدانی بدن ناشی میشود. بهطور طبیعی، سلولها در طول فعالیتهای معمول خود گونههای فعال اکسیژن تولید میکنند، اما این مولکولها معمولاً توسط سازوکارهای سمزدایی درون بدن حذف میشوند.
تولید بیش از رادیکالهای آزاد و شروع استرس اکسیداتیو
استرس گرمایی تعادل رادیکالهای آزاد و آنتی اکسیدانها را از دو طریق برهم میزند: یا تولید ROS (رادیکالهای آزاد) را افزایش میدهد یا اثربخشی خط اول دفاعی بدن یعنی آنزیمهای کاتالاز (CAT)، گلوتاتیون پراکسیداز (GPx) و سوپراکسید دیسموتاز (SOD) را کاهش میدهد. آنزیم SOD نقش محوری دارد زیرا تبدیل رادیکالهای سوپراکسید را به پراکسید هیدروژن و اکسیژن کاتالیز میکند. در ادامه، آنزیم کاتالاز (CAT) پراکسید هیدروژن را به آب و اکسیژن بیضرر تبدیل میکند. در حالیکه GPx با استفاده از گلوتاتیون، هیدروپراکسیدهای آلی را به الکلهای متناظر احیا مینماید. هنگامیکه بهدلیل قرارگیری طولانیمدت در معرض گرما، ROS در بدن تجمع بیش از حد پیدا میکند، این مولکولها اجزای حیاتی سلول مانند پروتئینها، لیپیدها و بهویژه DNA را تخریب میکنند. شدت استرس اکسیداتیو القاشده توسط گرما به عواملی مانند شدت و مدت زمان مواجهه بستگی دارد.
اگرچه در آغاز استرس گرمایی سطوح آنزیمهای آنتیاکسیدانی ممکن است افزایش یابد، اما قرارگیری طولانیمدت منجر به کاهش مزمن این آنزیمها میشود که آثار مخرب استرس اکسیداتیو بر سلولهای بدن را تشدید میکند. استرس گرمایی سطوح mRNA گرلین و کولهسیستوکینین را تغییر میدهد، که با محور HPA مرتبط است و اشتها را سرکوب میکند.
اختلال در تعادل اسید-باز
یکی دیگر از تغییرات فیزیولوژیکی مرتبط با استرس گرمایی، اختلال در تعادل اسید-باز بدن است. در اثر نفسنفس زدن، پرندگان نرخ تنفس خود و خنکسازی تبخیری ریهها را افزایش میدهند. این وضعیت معمولاً موجب میشود که دفع دیاکسیدکربن از طریق دستگاه تنفسی افزایش یابد و حتی از میزان تولید سلولی دیاکسیدکربن نیز فراتر رود. در نتیجه، سامانه بافری بیکربنات طبیعی خون دستخوش تغییر میشود. به طور مشخص، سطوح اسید کربنیک و یونهای هیدروژن کاهش مییابد در حالی که سطح یون بیکربنات در حد بالایی باقی میماند. این پدیده که بهعنوان آلکالوز تنفسی (Respiratory alkalosis) شناخته میشود، تأثیر عمیقی بر عملکرد کلیوی دارد. عدم تعادل در وضعیت اسید-باز خون با کاهش عملکرد در جوجههایگوشتی و مرغهای تخمگذار مرتبط دانسته شده است. اگرچه کاهش عملکرد حاصل عوامل متعددی است، اما یکی از مهمترین عوامل آن میزان خوراک مصرفی میباشد.
کاهش مصرف خوراک بر اثر تغییرات فیزیولوژیک
توضیح فیزیولوژیکی کاهش مصرف خوراک در دوره استرس گرمایی به تغییرات هورمونی نسبت داده میشود. محور نورونی-هیپوتالاموسی نقش کلیدی در تنظیم مصرف خوراک و تعادل انرژی بدن ایفا میکند. هورمونهای مختلفی از طریق محور نورونی-هیپوتالاموسی در حفظ مصرف خوراک و تعادل انرژی نقش دارند. مطالعات پیشین نشان دادهاند که در طیور، گرلین و کولهسیستوکینین نخستین هورمونهایی هستند که در تنظیم اشتها نقش دارند. سطوح mRNA و غلظتهای گرلین و کولهسیستوکینین در خون و بخشهای اصلی دستگاه گوارش جوجهگوشتیها و مرغهای تخمگذار، پس از قرارگیری در معرض استرس گرمایی بهطور معناداری تغییر مییابند. این تغییرات هورمونی همچنین ارتباط نزدیکی با سطوح هورمون CRH دارند.
قرارگیری طولانیمدت در معرض استرس موجب فعالسازی محور هیپوتالاموس–هیپوفیز–آدرنال (HPA) شده و در نتیجه ترشح هورمون CRH از هیپوتالاموس را تحریک میکند. این روند یک آبشار پاسخهای هورمونی را آغاز میکند که در آن غده هیپوفیز تحریک به ترشح هورمون ACTH میشود. در ادامه، ACTH تولید و ترشح هورمونهای کورتیکواستروئیدی توسط غدد فوقکلیوی (آدرنال) را تحریک مینماید. بنابراین، توضیح داده میشود که چرا کورتیکواستروئیدها، بهویژه کورتیکوسترون، بهعنوان شاخصهای زیستی (Biomarkers) استرس گرمایی در طیور مورد استفاده قرار گرفتهاند.
تأثیر استرس گرمایی بر ایمنی و سلامت روده
تأثیر استرس گرمایی بر سامانه درونریز (اندوکرین) منجر به سرکوب ایمنی میشود که پیامد مستقیم این دگرگونی فیزیولوژیکی است. پاسخ ایمنی بهطور دقیق توسط دستگاه عصبی مرکزی از طریق شبکهای پیچیده شامل دستگاههای عصبی، درونریز و ایمنی تنظیم میگردد. در سالهای اخیر، پژوهشهای گستردهای اثرات زیانبار استرس گرمایی را بر طیور بررسی کردهاند و بهطور مستمر کاهش در کارکردهای ایمنی را نشان دادهاند. بهویژه، قرارگیری مداوم در دماهای محیطی بالا با کاهش شاخص اندامهای لنفاوی مرتبط دانسته شده است که نشانگر سامانه ایمنی تضعیفشده میباشد.
علاوه بر این، وزن نسبی جگر در مرغهای تخمگذارِ دچار استرس گرمایی، کاهشیافته و نیز کاهش شدید آنتیبادیهای در گردش، بهویژه IgM و IgA، در جوجههای گوشتی که در معرض دماهای زیستمحیطی بالاتر از محدوده توصیهشده بودهاند، مشاهده شده است. در مورد پاسخ ایمنی سلولی، پژوهشگران فعالیت فاگوسیتی (بیگانهخواری) ماکروفاژها را کاهشیافته، شمار لنفوسیتهای T را کمتر و میزان بیان اینترلوکینها را در جوجهگوشتیهای مواجه با چالش استرس گرمایی کاهشیافته گزارش کردهاند. سطح بالاترِ کورتیکواستروئیدها و کاتکولآمینها که طی آسیب گرمایی مشاهده میشود میتواند تکثیر سلولهای ایمنی و تولید آنتیبادیها را مختل سازد که این امر میتواند توضیحدهنده کاهش اندازه اندامهای لنفاوی باشد. کاهش IgM و IgA با تضعیف میکروبیوم مرتبط است، که عفونتها را افزایش میدهد.
2) استرس سرمایی در طیور
استرس سرمایی در طیور میتواند در نتیجه قرار گرفتن در معرض دماهایی پایینتر از محدوده توصیهشده برای پرورش رخ دهد. برخلاف استرس گرمایی که پدیدهای بهخوبی مستندسازی شده است، استرس سرمایی در متون علمی کنونی کمتر رواج دارد. در واقع، بهدلیل افزایش جهانی دمای کره زمین، گرایش پژوهشهای فعلی معطوف به افزایش مقاومت پرندگان در برابر این دمای روبهافزایش است. با این حال، باید اذعان داشت که استرس سرمایی میتواند آثار زیانباری بر رفاه، رشد و رفتار طیور بر جای گذارد. بهویژه، در برخی کشورها یک عامل تنشزای زیستمحیطی مهم بهشمار میآید. پژوهشهای ۲۰۲۴ نشان میدهند که استرس سرمایی در زمستانهای سرد اروپا، تولید را تا ۱۵% کاهش میدهد.
پاسخهای فیزیولوژیکی و ایمنیشناختی به استرس سرمایی
جوجههای تازه از تخم درآمده بهویژه در روزهای پس از تفریخ بسیار در برابر استرس سرمایی آسیبپذیرند، زیرا توانایی محدودی در تنظیم دمای بدن خود دارند. اگرچه این کمبود حدودیک هفته پس از بیرونآمدن از تخم بهطور چشمگیری اصلاح میشود اما سازگاری با سرما تنها تا حد محدودی میتواند رخ دهد. پژوهشهای پیشین ثابت کردهاند که قرار گرفتن در معرض دمای پایین محیط در سنین کم میتواند اثری شدید بر فیزیولوژی جوجهها داشته باشد. برای مثال، قرار دادن جوجهگوشتیها در دمای ۴ درجه سلسیوس بهمدت ۲۴ ساعت موجب تغییر در برخی ژنهای مرتبط با متابولیسم چربی در هیپوفیز شده است و نشان میدهد که استرس سرمایی قادر است بر متابولیسم چربی اثر بگذارد. علاوه بر این، آشکار شده است که قرار دادن جوجهها در معرض استرس سرمایی، تولید حرارت بدن را تغییر داده و بیان ژنهایی مانند avUCP، AMPKα و PPARα را افزایش میدهد که بهترتیب با تولید حرارت و متابولیسم اسیدهای چرب ارتباط دارند. سایر تغییرات فیزیولوژیکی گزارششده شامل کاهش مصرف اکسیژن، کاهش دمای بدن، کاهش نرخ تنفس و کاهش اتلاف آب تنفسی هستند.
کاهش دریافت اکسیژن در دورههای سرمایی میتواند فشار جزئی اکسیژن در خون پرندگان را کاهش دهد. علاوه بر این، دماهای پایین هوا موجب افزایش نیاز به اکسیژن، بالا رفتن جریان خون و افزایش برونده قلبی میشوند که این امر باعث افزایش فشار شریان ریوی شده و بار مضاعفی بر بطن راست قلب وارد میکند. استرس سرمایی همچنین نیاز متابولیکی به اکسیژن را بیشتر تشدید میکند.
بهطور گسترده پذیرفته شده است که انواع تنشزاهای غیربیولوژیکی میتوانند شدت بیماریهای عفونی را افزایش دهند. بنابراین، احتمالاً استرس سرمایی نیز منجر به نتایجی مشابه میشود، گرچه سازوکارهای زیربنایی آن هنوز بهطور کامل روشن نشدهاند. پیشنهاد شده است که استرس سرمایی میتواند پاسخ ایمنی پرندگان را از طریق تأثیر همزمان بر مسیرهای زیستانرژیکی و درونریز (اندوکرینی) تغییر دهد. در واقع، در طیور باور بر این است که کارکردهای ایمنی و ترموتنظیمی هر دو از منابع انرژی مشترکی تغذیه میشوند. از اینرو، در طول مواجهه با سرما، افزایش نیاز انرژی برای تنظیم حرارت بدن ممکن است انرژی در دسترسِ لازم برای عملکرد مناسب سامانه ایمنی را محدود کند. این امر میتواند منجر به کاهش تولید سلولهای ایمنی و تضعیف واکنشهای ایمنی گردد. با این حال، آثار استرس سرمایی بر پاسخهای ایمنی خاص بهنظر ناهمخوان و متناقض است.
بررسی برخی از مطالعات در رابطه با تأثیر استرس سرمایی بر عملکرد سیستم ایمنی
برای نمونه برخی از محققین کاهش در پاسخهای آنتیبادی در مرغهای تخمگذار تکقفسی پس از قرارگیری در معرض استرس سرمایی را گزارش کردند. با این حال، چنین کاهشهایی در مرغهایی که در قفسهای گروهی نگهداری میشدند مشاهده نگردید. از سوی دیگر در یک آزمایش محققان هیچ نشانهای از کاهش کارایی ایمنی هومورال در بلدرچین شمالی پس از القای آزمایشی استرس سرمایی مشاهده نکردند. برعکس، مواجهه با سرما اثر منفی بر توان ایمنی هومورال در پرندگان بلوتیت (Blue tit) و نیز در مرغهای تخمگذار داشته است. استرس سرمایی همچنین سبب سرکوب ایمنی سلولی در خروسهای جوان گردید، درحالیکه جوجهمرغهای تخمگذار در حال رشد افزایش فعالیت ایمنی سلولی در اثر استرس سرمایی را نشان دادند. این تناقضات وابسته به نژاد و شرایط گروهی است (Ncho et al., ۲۰۲۴).
تأثیرات استرس سرمایی بر عملکردهای تولیدی
تأثیرات دماهای پایین محیطی بر عملکرد تولیدی پرندگان قابلتوجه و گسترده است، فارغ از آنکه برای تولید گوشت یا تخممرغ باشند. برای نمونه، قرار گرفتن طولانیمدت در معرض استرس سرمایی بهمدت ۷۲ ساعت میتواند باعث کاهش شدید رشد و افزایش ضریب تبدیل خوراک شود. به همین ترتیب، اثرات منفی تماس با سرما زمانی تشدید میشوند که جوجههای گوشتی بهمدت چند روز در معرض استرس سرمایی قرار گیرند.
مدت زمان تماس، عاملی بسیار مهم در شرایط استرس سرمایی است. کاهشهای مشاهدهشده در رشد طی استرس سرمایی احتمالاً با افزایش مصرف انرژی موردنیاز برای حفظ دمای بدن پرنده (homoeothermy) مرتبط است. به همینگونه، مرغهای تخمگذار برای تولید تخممرغ به انرژی مصرفی بالا و مقادیر زیاد مواد معدنی نیاز دارند و مشخص شده است که دماهای پایین موجب افزایش مصرف خوراک روزانه در عین کاهش تولید تخممرغ در مرغهای تخمگذار صنعتی میشوند، که در نهایت بازده خوراک را کاهش میدهد. مطالعات دیگری نیز کاهش همزمان وزن بدن و افت تولید تخممرغ را در مرغهای بَدَوی (Bedouin hens) پس از مواجهه با استرس سرمایی گزارش کردهاند.
اگرچه ثابت شده است که دماهای حدی بر عملکرد تولیدی اثر میگذارند، بهنظر میرسد اثرات استرس سرمایی با وجود رابطهای معکوس میان مصرف خوراک و خروجی تولید مشخص میگردد. بنابراین، برای ارزیابی رفتار تغذیهای پرندگان در شرایط دمای پایین و بهمنظور گسترش ادبیات علمی محدود فعلی پیرامون این موضوع، به پژوهشهای بیشتری نیاز است.
جدول ۱: مقایسه اثرات استرس گرمایی و سرمایی بر پارامترهای تولیدی (جدید و گسترشیافته)
|
منبع |
پارامتر |
استرس گرمایی |
استرس سرمایی |
|
Ncho et al., ۲۰۲۴ |
رشد و زن |
کاهش ۲۰-۳۰% (به دلیل سرکوب اشتها) |
کاهش ۱۵-۲۵% (به دلیل مصرف انرژی حرارتی) |
|
Zhou et al., ۲۰۲۱ (بهروزرسانی ۲۰۲۵) |
ضریب تبدیل خوراک (FCR) |
افزایش ۰.۲-۰.۵ |
افزایش ۰.۱-۰.۳ |
|
Iraqi et al., ۲۰۲۴ |
ایمنی (IgA/IgM) |
کاهش شدید (سرکوب HPA) |
متناقض (کاهش در هومورال، افزایش سلولی در برخی نژادها) |
3) استرس صوتی
صدا (Noise) بهعنوان صوت ناخواسته یا مزاحمی تعریف میشود که میتواند پیوسته یا گاهبهگاه باشد. از دیدگاه زیباشناختی، صدا از موسیقی متمایز است؛ اولی غیرعمدی و ناخوشایند است در حالیکه دومی گوشنواز و دلپذیر است. صدا منبع بالقوهای از استرس برای حیوانات محسوب میشود. همانگونه که در طیف گستردهای از مطالعات گزارش شده است. بهویژه، نشان داده شده است که صدا، پاسخ ترس (fear response) را در جانوران برمیانگیزد و بهعنوان یک عامل استرسزا در دام و طیور بهثبت رسیده است. عوامل گوناگونی در تعیین اثر صدا بر بهرهوری و رفتار حیوانات نقش دارند. با این حال، عوامل ذاتیِ مربوط به خودِ حیوان، مانند تواناییهای شنواییِ گونهها و نژادهای مختلف و بهویژه سن و وضعیت فیزیولوژیکی حیوان در هنگام تماس، نقش حیاتی در تعیین نوع پاسخهای برانگیخته شده ایفا میکنند.
ویژگیهای صدا همچون شدت (dB)، فرکانس (Hz)، مدت زمان و قابلیت ایجاد ارتعاش از عوامل مهمی هستند که هنگام بررسی آثار زیانبار صدا بر حیوانات باید مورد توجه قرار گیرند. صدا دارای قابلیت ایجاد ارتعاش است، ازاینرو پژوهشگران را بر آن داشته تا بررسی جامعی از اثرات نامطلوب آن بر طیور انجام دهند.
در یک مطالعه اولیه مشخص شد که در بازه فرکانسی ۱ تا ۱۰ کیلوهرتز، پرندگان در مقایسه با انسانها حساسیت شنوایی بیشتری دارند. همچنین، پرندگان تواناییهای پیشرفتهای در پردازش سیگنالهای صوتی از خود نشان داده و نسبت به محرکهای صوتیِ فرکانس پایین حساس بودند. در واقع، جنین مرغ از روز دوازدهم دوره جوجهکشی، شروع به دریافت سیگنالهای درونی حلزونی میکند. این دوره به عنوان مرحله پیششنوایی پرندگان نامگذاری شده است. از روز شانزدهم جوجهکشی، حلزون گوش توانایی تشخیص یا رمزگذاری صدا را پدیدار میکند. بهتدریج با رشد جوجهها، حساسیت آنان به صداهای فرکانس بالا افزایش مییابد.
در هر واحد دامداری یا مرغداری، دو منبع اصلی تولید صدا میتواند وجود داشته باشد. یکی از این منابع صدا میتواند دستگاهها و تجهیزات گوناگون مورد استفاده در واحد باشد، درحالیکه منبع دیگر از خود حیوانات ناشی میشود. دستگاههای متعدد درگیر در مدیریت روزمره مزارع مانند خوراکپاشها، تهویهها و فنها و همچنین تجهیزاتی که کارگران بهکار میبرند (همچون چرخبرها، شستوشوگرها و حتی مکالمات پرسنل) نمونههایی از منابع صدا در سالنهای نگهداری حیوانات هستند. حرکت حیوانات بزرگ، که اغلب با زنجیرهای فلزی مهارشدهاند، میتواند در حین جابجایی صداهای زیادی ایجاد کند. افزون بر این، فعالیتهایی همچون جویدن، گازگرفتن، تغییر وضعیت بدن و تولید اصوات توسط حیوانات نیز به محیط صوتی مزرعه میافزایند.
در سالهای ۲۰۰۸ تا ۲۰۱۰، مطالعات انجامشده در مزارع طیور شامل مرغهای تخمگذار، جوجه گوشتی و پولت نشان داد که خوراکرسانها و توزیعکنندگان خوراک اصلیترین منابع صدا در محیط مرغداری هستند. مطالعه دیگری نشان داد که جوجهگوشتیها در طول فرآیند کشتارگاهی در معرض صداهای بهشدت بلند قرار میگیرند. اگرچه اثرات صدا بر طیور بهطور کامل بررسی نشده است، اما مطالعات نشان دادهاند که میتواند بهطور منفی بر رفتار و عملکرد تولیدی پرندگان تأثیر بگذارد. صداهای >۹۰ dB با تغییرات میکروبیوتا مرتبط است، که تنوع باکتریایی را کاهش میدهد و ایمنی را تضعیف میکند.
تأثیر سروصدا بر تولید و رفتار طیور
سروصدا میتواند به طور منفی بر عملکرد تولیدی و رفتار پرندگان تأثیر بگذارد. در یک گزارش شدت صدای ۱۱۵ دسیبل باعث وقفه در عملیات جوجهکشی مرغها شد. با این حال، در همان مطالعه، شدت صدای ۹۶ دسیبل تأثیری بر میزان جوجهدرآوری یا کیفیت جوجههای تولیدشده نداشت. در آزمایش دیگری، پژوهشگران مشاهده کردند که جوجههایی که در معرض سروصدایی با شدت ۸۰ تا ۱۱۸ دسیبل قرار گرفتند، هیچگونه تأثیری بر رشد نشان ندادند، اما آنهایی که در معرض صداهای ۱۰۰ تا ۱۱۸ دسیبل بودند، رفتار تجمعی (ازدحام) از خود نشان دادند.
بررسی مطالعات محققین
برخی از محققین گزارش کردند که سروصدا اثر نامطلوبی بر افزایش وزن بدن، مصرف خوراک یا ویژگیهای رفتاری جوجههای گوشتی نداشت. با این حال گروهی دیگر از محققین دریافتند که قرارگیری در معرض صدا به میزان ۱۰۰ دسیبل باعث کاهش تعداد تلاشها برای القای بیحرکتی تونیک (TI) در جوجههای گوشتی شد. یکی از نخستین مطالعات بررسیکننده تأثیر سروصدا بر مرغهای تخمگذار انجام شد و نشان داد قرارگیری کوتاهمدت در معرض صدای هواپیما تأثیر منفی بر تولید تخممرغ ندارد. با این حال، دورههای طولانیمدت استرس ناشی از صدا منجر به کاهش تولید تخممرغ گردید. همچنین در آن مطالعه نشان داده شد که مدتزمان بیحرکتی تونیک (TI) در مرغهایی که بهمدت ۶۰ دقیقه در معرض صدای با شدت ۹۰ دسیبل قرار داشتند بهطور قابل توجهی طولانیتر بود.
از سوی دیگر محققان یافتند که صدا تأثیری بر بهرهوری مرغهای تخمگذار ندارد، در حالی که لرزشهایی بیش از ۱٫۰ سانتیمتر بر ثانیه بر بهرهوری در مرغهای تخمگذار تأثیرگذار بود. با این حال، عادتپذیری بوقلمونها به صدای شبیهسازیشده و واقعی پرواز هیچ تفاوتی در اثرگذاری بر رشد نشان نداد! در مطالعه دیگری که توسط Woolf و همکاران (۱۹۷۶) بر روی بلدرچین ژاپنی انجام شد، مشخص گردید که قرارگیری در معرض تحریک شنوایی با شدت ۸۰ دسیبل بهمدت ۲ ساعت در سه روز پایانی دوره انکوباسیون، روند خروج جوجهها از تخم را تسریع نمود.
بهطور معمول، ماکیان اهلی هنگام مواجهه با صداهای بلند و ناگهانی، با چرخش سر و یک واکنش کوتاهمدت وحشتزدگی پاسخ میدهند. با این حال، پرندگان توانایی قابلتوجهی در عادتپذیری سریع دارند و ظرف چند دقیقه پس از قطع تحریک صوتی، به فعالیتهای طبیعی خود بازمیگردند. واکنش ترس در مرغها و پرندگانی که بهمدت ۱ ساعت در معرض صدای ۹۰ دسیبل ناشی از کامیونها، قطارها و هواپیماها قرار گرفتند، نسبت به مرغهایی که در محیطهایی با صدای ۶۵ دسیبل عمدتاً ناشی از آواز طبیعی نگهداری میشدند، افزایش یافت.
در مرغها، قرارگیری در معرض محرکهای صوتی با شدت ۹۵ دسیبل در فرکانس ۵۰۰ هرتز، واکنشی از وحشت ایجاد کرد که شامل دوره نهفتگی، دویدن، بیحرکتی کامل، حرکات تند و کوچک سر و رفتار خوابآلودگی بود. پرورش طیور در محیطی با حداقل وقوع صداهای بلند و ناگهانی بسیار حیاتی است، زیرا چنین صداهایی سبب بروز رفتارهای هیستریک (وحشت جمعی) در نژادهای مختلف مرغ میگردد.
در ۱۰ مورد از ۲۱ مطالعه، گلههای مرغ بالغ حدوداً ۳۵ هفتهای هنگام قرارگیری در معرض صدای ضبطشده ماشینآلات و آواهای پرندگان، رفتار نوکزدن به پر (feather-pecking) از خود نشان دادند. در گونههای دیگر طیور مانند بوقلمونهای وحشی هیچ رفتار غیرعادی در مواجهه با صداهای مهیب گزارش نکردند، در حالی که بروز وحشت و پرخاشگری را در بوقلمونهای در معرض صدا مشاهده کردند.
تأثیر سروصدا بر هماتولوژی طیور
تأثیر سروصدا بر هماتولوژی طیور قابلتوجه و زیانآور است. قرارگیری طولانیمدت در معرض صدای شدید با افزایش فعالیت دستگاه عصبی خودمختار مرتبط است. این فعالسازی پایدار با افزایش فعالیت محور هیپوتالاموس–هیپوفیز-آدرنال، افزایش نرخ متابولیک، فشار خون بالا و تاکیکاردی همراه است. بهخوبی مشخص است که محور HPA بخش کلیدی از دستگاه درونریز است و در حفظ هموستاز نقش اساسی دارد. این محور شامل سه جزء اصلی است:
- هیپوتالاموس که هورمونهایی همچون دوپامین و CRH (هورمون آزادکننده کورتیکوتروپین) را سنتز میکند.
- غده هیپوفیز، بهویژه لوب قدامی آن، که هورمون ACTH را ترشح میکند.
- و غده آدرنال.
ترشح زنجیرهای هورمونها با فعالسازی هیپوتالاموس آغاز میشود که CRH را ترشح میکند و متعاقباً غده هیپوفیز را برای آزادسازی ACTH تحریک مینماید.
غده آدرنال نیز به دو بخش تقسیم میشود:
مدولای آدرنال که مسئول ترشح کاتکولآمینهایی مانند اپینفرین و نوراپینفرین است و قشر آدرنال که هورمونهای استروئیدی شامل کورتیزول، کورتیکوسترون و آلدوسترون را تولید میکند.
قشر آدرنال با تحریک ACTH برای ترشح گلوکوکورتیکوئیدها فعال میشود. کورتیکوسترون هورمون اصلی استرس در پرندگان است. تولید این قبیل هورمونها در بدن هنگام پاسخ به استرس بهطور چندبرابر افزایش مییابد. افزایش ناگهانی سطح این هورمونها تغییرات متابولیکی درون سلولی ایجاد میکند و فرآیندهایی مانند لیپولیز و پروتئولیز را آغاز مینماید که میتواند منجر به افزایش سطح گلوکز خون شود. در دورههای طولانیمدت استرس، بالا بودن سطح خونی گلوکوکورتیکوئیدها به بروز اختلالاتی منجر میشود که باعث کاهش وزن اندامهایی نظیر غدد لنفاوی و طحال میگردد؛ در نتیجه، عملکرد ایمنی حیوان تضعیف شده و حساسیت آن به عوامل بیماریزا افزایش مییابد.
پاتوفیزیولوژی استرس ناشی از صدا به صورت خلاصه در شکل ۳ نمایش داده شده است.

در مطالعهای توسط Borg (۱۹۸۱) مشاهده شد که قرارگیری در معرض صدا باعث افزایش تدریجی در سطح پایه استروئیدها در مرغهای نژاد "وایت لگهورن" شد. با این حال، McFarlane، Curtis و Simon یافتند که قرارگیری مداوم جوجههای گوشتی در معرض صدا با شدت ۸۰ یا ۹۰ دسیبل تأثیر معنیداری بر نسبت هتروفیل به لنفوسیت یا سطح پلاسمایی کورتیکوسترون نداشت. با این وجود، همین نویسندگان همچنین گزارش کردند که درصد مونوسیتها و شدت ضایعات دوازدهه ناشی از کوکسیدیوز در پرندگانی که در معرض صدای مداوم با شدت ۸۰ یا ۹۵ دسیبل قرار داشتند، افزایش یافت. علاوه بر این، افزایش قابلتوجهی در گرانولوسیتهای بازوفیل زمانی مشاهده شد که جوجههای گوشتی در معرض شدت صدای ۷۰ و ۸۰ دسیبل قرار گرفتند.
Gross (۱۹۹۰) گزارش کرد که قرارگیری در معرض صدای ۱۰۴ دسیبل بهمدت ۳۰ ثانیه، نسبت هتروفیل به لنفوسیت را در مرغها افزایش داد. Campo و همکاران (۲۰۰۵) نیز گزارش کردند که مرغهای تخمگذار در معرض صدا، دارای نسبت هتروفیل به لنفوسیت بهطور معنیداری بالاتر بودند. Chloupek و همکاران (۲۰۰۹) یافتند که جوجههای گوشتی قرارگرفته در معرض شدت صدای ۸۰ و ۱۰۰ دسیبل دارای سطح پلاسمایی کورتیکوسترون بهطور معنیداری بالاتر بودند، اگرچه قرارگیری در معرض این محرکهای صوتی تأثیری بر سطح گلوکز و غلظت تریگلیسرید نداشت. با این حال، قرارگیری در معرض محرکهای صوتی با شدت ۱۰۰ دسیبل باعث افزایش معنیدار در سطح کلسترول و تریگلیسرید شد. علاوه بر این، طیور در هنگام حملونقل در واکنش به صدا، افزایش ضربان قلب و نرخ تنفس، همراه با ترشح بیشتر هورمونهای استرس از خود نشان دادند. مطالعات ۲۰۲۴ بر اثرات صوتی بر میکروبیوم تأکید دارند، که التهاب روده را افزایش میدهد.
4) استرس ارتعاشی در تولید طیور
هر حرکت تکرارشوندهای که در فواصل منظم رخ میدهد، ارتعاش شناخته میشود. ارتعاشات مکانیکی جزئی ذاتی از فرآیند پرورش طیور هستند که از مرحله جوجهکشی آغاز شده و در تمام مراحل تولید ادامه مییابند. نقش اصلی استرس ارتعاشی در رفتار و فیزیولوژیِ حیوانات در دوره حملونقل آشکار میشود. بیماریِ حرکتی (motion sickness) و احساس ناراحتی، پیامدهای نامطلوب شناختهشدهای هستند که بر اثر حرکت وسایل نقلیه ایجاد میشوند.
در طی حملونقل، بهویژه در مسیرهای ناهموار، حیوانات در معرض انتقال قابلِتوجه ارتعاشات کف وسیله نقلیه قرار میگیرند. این جابجایی مرکز ثقل، تعادل بدن آنها را مختل میسازد و در پی آن احساس ناراحتی، اختلال در رفاه و در نهایت، کاهش کیفیت گوشت رخ میدهد. عوامل ذاتیِ گوناگونی همچون وزن بدن، اندازه، سن، جنس، سطح سلامت بدنی و وضعیت قرارگیری بدن، پاسخ حیوان به استرس ارتعاشی را تعیین میکنند. مشخص شده است که موجودات زنده در برابر ارتعاشاتِ وارده پاسخهای متفاوت از خود نشان میدهند.
بیشینه سطح ارتعاش در فرکانس رزونانس رخ میدهد؛ جایی که بدن بیشترین انرژی مکانیکی ممکن را در مقایسه با هر فرکانسِ دیگر جذب میکند. در برخورد با ارتعاشات، اعضای بدن بهصورتی یک سامانه مکانیکیِ ناهمگن عمل میکنند که در نتیجه منجر به جابجاییِ ارگانهای داخلی میشود. مطالعات نشان دادهاند که فرکانسهای اساسیِ کفِ لوازم یا ادوات حمل پرندگان معمولاً بین ۱ تا ۴ هرتز متغیر است و یک اوج دیگر در بازه ۱۰ تا ۱۲ هرتز دیده میشود. این فرکانسها بهطور نزدیکی با فرکانسهای اثرگذار بر جوجهگوشتیِ دو کیلوگرمی منطبق بوده و احتمالاً میتوانند پاسخهای استرس ارتعاشی را برانگیزند. افزون بر آن، قرارگیریِ مزارعِ پرورش دام و طیور در نزدیکیِ کارگاهها، خطوط راهآهن، محوطههای ساختمانی یا واحدهای مشابه، میتواند آنها را در معرض استرسِ ناشی از ارتعاشاتِ منتقلشونده از زمین قرار دهد.
همچنین منابع گوناگونی در ایجاد ارتعاش سهم دارند، از جمله هودهای تهویه، فنهای خروجی و انواع گوناگونِ تجهیزاتِ موتور دار. این ارتعاشات زنجیره تولید طیور را بهطور قابلِتوجهی مختل میکنند. اگرچه دانش موجود درباره فیزیولوژیِ استرس ارتعاشی اندک است، اما این پدیده با استرس صوتی ارتباط نزدیکی دارد، زیرا هر دو به صورت امواج انرژی عمل میکنند.
این پدیده یعنی استرس ارتعاشی بر کیفیت گوشت جوجهگوشتی در طیِ انتقال به کشتارگاه تأثیر میگذارد.
تأثیر ارتعاش بر تخممرغها
از دست رفتن تخممرغهای خوراکی یا لقاحیافته در اثر ترکها، شکستگیها و تغییر در کیفیت محتویات داخلی تخم مرغها، از جمله اثراتِ مشهود ارتعاش بر تخممرغهای طیور بهشمار میرود. قرار گرفتن تخممرغها در معرض ارتعاشات مکانیکی میتواند باعث تغییر در یکپارچگیِ پوسته، زرده و سفیده شود و به هدررفت، ایجاد ترک در پوسته، مایع شدن سفیده، کاهش مقادیر شاخصها و افت مقاومتِ غشای زرده بیانجامد. این تغییرات نشاندهندهٔ اثرات زیانآور ارتعاشات بر کیفیت درونی تخممرغها هستند.
بررسی برخی از مطالعات
در یک مطالعه پیشگام در گذشته که بر تخممرغهای لقاحیافته متمرکز بود، قرار دادن آنها در معرض ارتعاشات روزانه با سرعتِ ۳۶۰۰ حرکت در دقیقه به مدت ۱۵ دقیقه پیش از دوره جوجهکشی، هیچ تأثیری بر درصد جوجهدرآوری نشان نداد. با این حال، Taggart و همکاران (۱۹۹۰) کاهشِ تا ۶۸ درصدی نرخ جوجهدرآوری را در تخمهایی که در معرض ارتعاش قرار داشتند گزارش کردند. افزون بر این، نویسندگان اذعان کردند که تخمهایی که در معرض ارتعاش قرار میگیرند ممکن است دچار شکستگی در زرده یا تغییر در ساختارهای اولیه دیسک لقاحیافته شوند که توسعه موفق رویان را مانع میگردد. Shannon و همکاران (۱۹۹۴) تأثیر ارتعاشاتِ مکانیکی با فرکانسهای ۵ تا ۵۰ هرتز و شتابهایی در دامنه ۰٫۰۹ تا ۴٫۹۳ متر بر ثانیه مربع بر تخممرغهای لقاحیافته بررسی کردند. آنها نرخ مرگومیرِ ۳۱٫۹ درصدی را گزارش کردند و تأکید داشتند که دامنه و فرکانسِ ارتعاش هر دو با کاهش موفقیت در جوجهدرآوری ارتباط مستقیم دارند. Torma و Kovachne (۲۰۱۲) در تخمهایی که در معرض ارتعاش قرار داشتند، کاهش نرخ جوجهدرآوری و افزایش بروز ناهنجاریهای جنینی را ثبت کردند.
شایان ذکر است که تخممرغها در زنجیره تولید بهطوردائم در فرآیندهایی مانند جمعآوری، بستهبندی و حملونقل در معرض ضربات مکانیکی قرار دارند. Nazar و همکاران (۲۰۱۳) نتیجه گرفتند که ارتعاشات مکانیکی به طور مستقیم بر تلفات پیش از ورود به مزرعه تأثیر میگذارند. سطوح بالای ارتعاش در جادههای آسفالته و خاکی میتواند منجر به ترک یا شکستگیِ تخممرغها شود. ارتعاشات مکانیکی سبب کاهشِ کیفیتِ جوجهدرآوری و در نتیجه، کاهش تعداد جوجههای تازه متولدشده و با کیفیت بالا گردید.
تأثیر ارتعاش در جوجههای گوشتی
حملونقلِ جوجههای گوشتی در مسافتهای طولانی میتواند موجب بروز استرس قابلتوجهی شود، که این تنش با افزایش فاصله میان مزارع و کشتارگاهها تشدید میگردد. Randall و همکاران (۱۹۹۳) و Carlisle (۱۹۹۸) گزارش کردند که ارتعاشات موجب ایجاد ترس و ناراحتی در پرندگان میشوند و در نتیجه اثرات منفی بر کیفیت گوشت بر جای میگذارند. آنها در شرایط عملی مشاهده کردند که در جریان حمل پرندگان با کامیون به کشتارگاه، ارتعاشاتی با فرکانس تا حدود ۱۰ هرتز در محور عمودی و تا حدود ۱۸ هرتز در محور جانبی رخ میدهند. از آنجا که اندامهای داخلی درون حفره بدن بهطور محکم ثابت نیستند، هنگام قرار گرفتن در معرض ارتعاش میتوانند دچار جابجایی نسبی شوند. این جابهجایی متناسب با جرم اندامهای داخلی است. وقتی اندامها در حالت تشدید قرار میگیرند، ممکن است آسیبهایی از پارگی خفیف سلولی تا نارسایی شدید عملکردی و خونریزی داخلی متحمل شوند. بنابراین، تشدید (رزونانس) اندامهای داخلی میتواند بهعنوان یک محرک آزاردهنده قابلتوجه برای پرندگان در هنگام مواجهه با ارتعاش عمل کند.
حملونقلِ جادهای در جوجههای گوشتی بهطور تجربی نشان داده است که پاسخهای استرسی را فعال میسازد که این امر با افزایش نسبتِ هتروفیل به لنفوسیت و سطوح بالاترِ آنزیمهای کیناز در پلاسما ثابت شده است. همچنین گزارش شده است که ارتعاشات در بازه ۲ تا ۱۰ هرتز موجب کاهش قابلتوجهی در گلیکوژن عضلانی پرندگان میشود. افزون بر این، Carlisle (۱۹۹۸) گزارش کرد که سطوح مختلف ارتعاش (۲، ۵ و ۱۰ هرتز) منجر به افزایش سطح کورتیزول در پرندگان شد. مطالعات نشان میدهند که استرسِ ناشی از ارتعاش باعث بروز تغییرات بیوشیمیایی میشود و در نتیجه غلظت انتقالدهندههای عصبی مغز افزایش مییابد. با این وجود، Garcia و همکاران (۲۰۰۹) گزارش کردند که در جوجههایگوشتی در معرض ارتعاشات شبیهسازیشده حملونقل، هیچ تغییری در دمای مقعدی و کاهش وزن مشاهده نشد. Abeyesinghe و همکاران بیان کردند که پرندگان تمایل به اجتناب از ارتعاش دارند زیرا این پدیده سبب ترس در میان آنها میشود.
از آنجا که ارتعاش بهعنوان محرک استرسزا توسط پرندگان درک میشود، تمایل دارد واکنشهای استرسی را در بدن برانگیزد. این موضوع میتواند توجیهی برای سطوح بالاتر کورتیزول در پرندگانی باشد که در معرض فرکانسهای متفاوت ۲، ۵ و ۱۰ هرتز قرار گرفتهاند. مطالعات همچنین نشان دادهاند که جوجههای تازه از تخم خارجشده بهصورت طبیعی صداهای هماهنگ را به صداهای ناهماهنگ ترجیح میدهند.
از دیدگاه زیستی، نقش ترس، هدایت پرندگان به دور از خطرات احتمالی است و بنابراین بهعنوان یک مکانیسم بقاء سودمند عمل میکند. با این حال، در شرایط خاص، اگر پرندگان نتوانند از محرک آزاردهنده بگریزند، ترس میتواند اثرات زیانبار ایجاد کند. این ویژگیِ ترسپذیری تحت تأثیر مجموعهای از عوامل ژنتیکی و اپیژنتیکی قرار دارد. تأثیر عوامل محیطی بهویژه در مراحل اولیه رشد بسیار حائز اهمیت است. میزان ترسپذیری بهطور چشمگیری از محرکهای بیرونی که در مراحل ابتدایی زندگی تجربه میشوند، تأثیر میپذیرد. از اینرو، پرورش طیور در محیطی پیچیده میتواند میزان ترسپذیری را کاهش دهد. غیرواقعی است که تصور کنیم نویزهای محیطی را میتوان بهطور کامل در شرایط پرورش طیور حذف کرد. افزون بر این، کنترل کیفیت جادهها و کامیونها در طول حملونقل همواره امکانپذیر نیست. در نتیجه، لازم است راهبردهایی بر پایه یافتههای علمیِ نوین طراحی و اجرا شوند تا بتوان ارتعاشات را در تمامی مراحل زنجیره صنعت طیور از پیشهچ تا مزرعه و از مزرعه تا کشتارگاه کاهش داد؛ این کار میتواند از طریق بهکارگیری مفاهیم مهندسی در طراحی جعبهها، سینیها و مواد جدید برای وسایل نقلیه انجام گیرد.
برای به حداقل رساندن استرس ناشی از صدا و ارتعاش، ضروری است که محل استقرار و شیوه عملکرد فنهای تهویه و تجهیزات تغذیه با دقت و برنامهریزی مهندسی انتخاب گردد. علاوه بر این، شرطیسازی و عادتدهی پرندگان میتواند به کاهش میزان ترسپذیری آنها کمک کند. روش «حساسیتزدایی نظاممند» که با موفقیت برای درمان فوبیا در انسانها بهکار رفته است، باید برای پرندگان نیز چه در فوبیاهای طبیعی و چه در انواع پاتولوژیک مورد ارزیابی و تأیید قرار گیرد.
برخی نژادهای مرغ بهویژه نسبت به صدا حساساند و ممکن است حتی به اصوات خفیف با وحشت واکنش نشان دهند که این مسئله میتواند موجب خسارتهای قابلتوجه شود. ازاینرو، حساسیت و بروز رفتارهای ترسآمیز میتواند تحت کنترل مجموعهای از عوامل ژنتیکی و اپیژنتیکی قرار گیرد. بررسی این نوع روابط، پیش از یافتن روشی برای کاهش استرسِ ناشی از ارتعاش، امری ضروری است. بنابراین، یک رویکرد جامع که سطح تحمل پرنده و همچنین مراحل مختلف تولید و حملونقل را در نظر گیرد، میتواند راهکارهای مؤثری برای کاهش اثرات زیانبار صدا و ارتعاش در صنعت پرورش طیور فراهم سازد.
تنشهای نوری
نور نقشی حیاتی در آغاز الگوهای ترشح هورمونهای گوناگون دارد، تأثیر قابل توجهی بر رشد، بلوغ و تولیدمثل در طیور میگذارد. مدیریت نامناسب نور میتواند منجر به ناهنجاریهای بینایی، بدشکلیهای اسکلتی و مشکلات گردش خون در کنار سایر نارساییها گردد. برای بهبود تولید و افزایش بهرهوری در طیور، پرداختن به جنبههای مدیریتیِ محیطی همچون شدت نور، مدت زمان تابش، طول موج و الگوی دورهنوری ضروری است، زیرا این عوامل تأثیر بسزایی بر فعالیتهای فیزیکی مرغها دارند. یافتههای اخیر نشان میدهند که طول موج نور، بر توانایی طیور در پاسخگویی به استرس، ترس و افسردگی در دوره آسایش آنها تأثیرگذار است (شکل ۴).

این قرارگیری در معرض نور میتواند بر نرخ جوجهدرآوری تأثیر بگذارد، بهویژه زمانی که جنینها در طول دوره انکوباسیون در معرض نور طبیعی خورشید قرار گیرند. نوردهی نامنظم میتواند با تأثیر بر هستههای سوپراکیاسماتیک واقع در هیپوتالاموس، موجب اختلال در ریتمهای شبانهروزی شود؛ در نتیجه، هماهنگی عملکردهای فیزیولوژیک و رفتاری در انسان و حیوان تحتتأثیر قرار میگیرد. علاوه بر این، پویاییهای گروهی مرتبط با ریتم روزانه مانند تشکیل گله در پرندگان و تغذیه همزمان در طیور اهلی، میتواند فعالیت نوسانی مغز را در باندهای آلفا و مو بهبود بخشیده و همزمانی بینمغزی را در سطح دوگانه افزایش دهد.
برخی محققین با استناد به مطالعاتی نتیجهگیری کردند که نواحی آناتومیکی مهمی را که بر عملکرد بینایی طیور اثرگذارند، مشخص کردهاند:
- شبکیه با واکنش به نور محیطی به رشد مناسب و رفتار طبیعی پرنده کمک میکند.
- غده پینهآل، مسئول ترشح هورمونهای ملاتونین و سروتونین است که تحرک، دمای بدن و فصول جفتگیری را تنظیم میکنند.
- هیپوتالاموس که بهطور مستقیم هورمون آزادکننده گنادوتروپین (GnRH)، تعادل داخلی (هموستازی) و ساختارهای فیزیولوژیک را بر اساس شدت نور کنترل میکند و در تراکمهای بالا به طول موجهای کوتاه نیاز دارد. بنابراین، رنگ نور حاصل از طول موجهای مختلف از جمله طیفهای نامرئی بر سلامت و رفاه طیور اثرگذار است و واکنشهای رفتاری، بهویژه رفتارهای مرتبط با ترس را تحت تأثیر قرار میدهد. همچنین لازم است سایر اثرات نور، همانگونه که در جدول ۲ (در ادامه مطالب) نشان داده شده است، مورد بررسی قرار گیرند.
شدت نوری یا روشنایی
شدت نور در سالنهای پرورش طیور و سایر محیطهای مصنوعی بر حسب واحد لوکس (Lux) یا کندلا بر متر مربع (cd m⁻²) اندازهگیری میشود و تأثیر قابلتوجهی بر رفتار مرغها دارد. بر اساس برنامه نوری استاندارد برای جوجهگوشتیها، در طول ۷ روز نخست (دوره اولی پسازهچ) شدت نوری پیوسته معادل ۲۰ لوکس حفظ میشود و بعد از آن برای روزهای باقیمانده پرورش کاهشیافته و در سطح کمنور ۳ تا ۵ لوکس تنظیم میگردد. منابع نور مصنوعی معمولاً نوری با جریان متناوب ۵۰ هرتز در ثانیه تولید میکنند، که باعث نوسانات در شدت نور میشود بهگونهای که لامپها در هر ثانیه ۱۰۰ مرتبه روشن و خاموش میگردند. استفاده از نور روشنتر میتواند موجب افزایش فعالیت مرغها و کاهش وزن بدن آنها شود. افزایش فعالیت بدنی میتواند بهطور بالقوه بروز اختلالات اسکلتی و متابولیکی را کاهش دهد. جوجههای گوشتی که در معرض نور طبیعی شدیدتر (۸۵٫۲ لوکس) قرار گرفتند، فعالیت بیشتری نشان دادند؛ بهطوریکه در طول هفتههای دوم تا ششم، افزایش در بسامد رفتارهایی مانند ایستادن، حرکت، تغذیه و نوکزدن به زمین مشاهده شد.
این مشاهده نشان میدهد که بینایی پرندگان در شرایط نور طبیعی تکامل یافته است، بهگونهای که آنها قادر به درک نور لرزان (flicker) در فرکانسهای بالاتر هستند. این توانایی به بینایی تتراکروماتیک (چهاررنگبینیِ) آنها نسبت داده شده است که بهصورت نظری به آنها اجازه میدهد تا دو برابر انسانها رنگهای بیشتری را تشخیص دهند. بنابراین، حساسیت پرندگان اهلی نسبت به لرزش نوری (flicker sensitivity) میتواند بهراحتی باعث شود از نزدیکشدن به اشیایی که موجب ناراحتی یا استرس میشوند خودداری کنند؛ بهویژه در محدوده فرکانسی ۳۹ تا ۷۱ هرتز و سطوح روشنایی بین ۱۰ تا ۱۰۰۰ کندلا بر متر مربع.
نرخ رشد جوجهگوشتیهای نژاد «راس» که در شدت نوری ۲۰ لوکس (با فعالیت رفتاری بیشتر) پرورش یافتند، کمتر از جوجههایی بود که در شدت نوری ۵ لوکس نگهداری شدند، بهطوریکه منجر به کاهش وزن چشم و کاهش افزایش وزن کلی گردید. با این حال، کاهش تدریجی شدت نور از ۲۵ لوکس در ۷ روز نخست به ۲ تا ۵ لوکس، میتواند روشی پیشگیرانه برای بهبود عملکرد جوجههای گوشتی در زمینه کاهش خودخواری و نوکزدن پرها باشد و همچنین منجر به کاهش رفتارهای ایستادن و راهرفتن گردد.
پژوهشهای دیگر نیز نشان دادهاند که نوسانات شدت و کیفیت نور طبیعی در طول روز میتواند با تأثیر بر ریتمهای شبانهروزی و فراروزی، رفتار پرندگان درون گله را همزمان (سینکرون) کند و احتمالاً بر سلامت پاها نیز تأثیر بگذارد. برای نمونه، جوجههایی که در شدت نوری ۲۰۰ لوکس نگهداری شدند، در تمام دوره نوری زمان بیشتری را صرف مرتبکردن پرها (preening) کرده و پرهای سالمتری داشتند، در مقایسه با آنهایی که در نور ۵ لوکس بودند. با این حال، این تفاوت در شدت نور تأثیری بر سلامت پاها نداشت.
در مرغهای تخمگذار مشخص شد که قرارگیری در معرض نور LED با شدت ۵ و ۱۰۰ لوکس تأثیری بر عملکرد فیزیولوژیک تولیدی یا کیفیت تخممرغ در نژاد های-لاین W-80 ندارد. با این حال، مشاهده گردید که قرارگیری در معرض نور از روز هفتم تا بیستوهشتم بر دمای کلوآک و مقعد، میزان خوراک مصرفی، وزن بدن و درصد آلبومین اثر میگذارد. بنابراین نتیجهگیری شد که شدت نور ۵ لوکس و ۲۸ روز دوره سازگاری، بهترین شرایط برای مرغهای تخمگذار است.
در مطالعهای دیگر مشاهده شد که شدت نور بین ۹۹ تا ۳۲۳ لوکس، با فاصله ۸۳٫۸ سانتیمتر بین لامپها و دانخوریها، در مرغهای نژاد Lohmann-Brown موجب بلوغ جنسی زودرستر میشود که در نتیجه باعث افزایش تولید تخممرغ اما کاهش وزن آن (۵۹٫۶ گرم بهازای هر پرنده) میگردد. علاوه بر این، وزن مخصوص تخممرغها در مرغهای تخمگذار های-لاین W-36 ارتباط مستقیمی با کیفیت تخممرغ دارد. این کیفیت را میتوان با قرار دادن در معرض نور LED به شدت ۲۸ لوکس در شب بهبود بخشید که ثابت شده است عملکرد تولیدی را نیز ارتقا میدهد. پژوهشهای اخیر ارتباط میان شدت نور و طول موج را روشن ساختهاند. همچنین، اثر روشنایی (Illuminance) که بر حسب لوکس اندازهگیری میشود، بر رشد و رفتار جوجهگوشتیها نیز بررسی گردید.
رژیم دوره نوری یا برنامههای نوردهی
فراتر از عوامل متداولی مانند دما، رطوبت، سرعت جریان هوا و تابش، دوره نوری محیطی بهویژه چرخه ۲۴ ساعته روشنایی و تاریکی اطلاعات حیاتی درباره تغییرات روزانه نور خورشید فراهم میآورد که برای حفظ و همزمانسازی هموستاز فیزیولوژیک و رفتاری ضروری است. بهطور قابلتوجهی گزارش شده است که یک دوره نوری طولانی (۲۰ ساعت روشنایی و ۴ ساعت تاریکی) در جوجهگوشتیهای نژاد راس ۳۰۸ موجب کاهش کیفیت گوشت میشود، در حالیکه دوره نوری تقریباً پیوسته (۲۳ ساعت روشنایی و ۱ ساعت تاریکی) باعث افزایش سطح استرس و ترس در پرندگان میگردد. علاوه بر این، دورههای نوری تقریباً پیوسته موجب اختلال در بیان ژنهای ساعت شبانهروزی و کاهش تنوع میکروبیوتای سکوم در مرغهای تخمگذار نژاد هایلاین براون میشوند؛ بهطوریکه ریتمهای شبانهروزی القاشده در مراحل اولیه زندگی با سلامت دستگاه گوارش ارتباط دارند. با توجه به این یافتهها، طراحی و اجرای یک برنامه نوری دقیق و هدفمند برای بیشینهسازی رشد و سلامت طیور امری ضروری است که در نهایت موجب مزایای اقتصادی میگردد. مدت زمانی که مرغها در معرض نور قرار میگیرند که «برنامه نوردهی» نیز نامیده میشود تأثیر زیادی بر نوع گوشت یا تخممرغ تولیدی دارد. انواع مختلف برنامههای نوردهی تأثیر چشمگیری بر عملکرد، ایمنی و ترشح هورمونهایی که رشد و تولیدمثل را کنترل میکنند دارند. بنابراین، تنظیم دوره نوری بهصورت هدفمند جهت افزایش رشد و بهبود عملکرد اقتصادی در عین تضمین سلامت پرندگان مورد استفاده قرار میگیرد. این پدیده از طریق آزمایشهایی که فعالیت حرکتی جوجههای گوشتی را تحت شرایط نور تقریباً پیوسته و دوره نوری ۱۶ ساعت روشنایی و ۸ ساعت تاریکی بررسی کردهاند، تأیید شده است. سطح متوسط فعالیت در فاز روشنایی با کاهش مدت زمان نوردهی افزایش یافت.
جوجههای گوشتی که در معرض نوردهی ۲۳ ساعت روشنایی و ۱ ساعت تاریکی قرار گرفتند، موفق به ایجاد الگوی فعالیت شبانهروزی نشدند؛ در حالیکه در پرندگان گروه ۸ ساعت تاریکی و ۱۶ ساعت روشنایی چنین الگویی بهخوبی شکل گرفت. این یافتهها بر اهمیت ریتمهای شبانهروزی بهعنوان شاخصی از رفاه در حیوانات اهلی تأکید دارند. پژوهشها نشان دادهاند که قرارگیری در معرض نور روشن (۴۰ لوکس؛ ۱۷ ساعت روشنایی در روز) مشکلات مربوط به سلامت پا و چشم را کاهش داده و تأثیر مثبتی بر لاشه سرد، گوشت سینه و بازده بالها دارد.
در مقابل، برنامههای نوری که شامل دورههای طولانیتر تاریکی (حدود ۷ ساعت در روز) هستند، با کاهش تعداد پرندگان دچار درد، رفاه جوجههای گوشتی را بهبود میبخشند. در جوجههای گوشتی، قرارگیری در معرض نور برای تحریک رشد اسکلتی از طریق افزایش مصرف خوراک حیاتی است و بر عملکرد کلی تولیدی تأثیر قابلتوجهی دارد. این فرآیند همچنین بر توانایی پاسخدهی جنینهای مرغ به محرکهای تهدیدکننده مانند شکارچیان تأثیر دارد که با پدیده بیحرکتی تونیک (Tonic immobility) سنجیده میشود.
در ارزیابی درجه ترس در پرندگان، شاخص بیحرکتی تونیک (TI) معیار مناسبی محسوب میشود. در یک مطالعه نشان داده شد که مرغهایی که در معرض ۲۳ ساعت روشنایی قرار گرفتند، در روز دهم مدت بیحرکتی تونیک طولانیتری داشتند، هرچند این روند در روز سیوششم معکوس گردید. پرندگانی که در معرض نور افزایشی (۲۴ ساعت روشنایی در سه روز نخست) بودند نیز در همان مقطع مدت TI طولانیتری از خود نشان دادند. این موضوع بیانگر آن است، پرندگانی که در معرض نوردهی منقطع (Interrupted Lighting) قرار دارند، ممکن است در سازگاری با تغییرات طول روز در مراحل بعدی زندگی دچار مشکل شوند و در نتیجه پاسخهای ترس شدیدتری نسبت از خود بروز دهند.
نوردهی پیوسته (۲۴ ساعت روشنایی) تأثیر منفی بر رشد اندامهای حرکتی جنین داشته و استحکام استخوانهای پا را در مراحل بعدی زندگی کاهش میدهد. در مقابل، اجرای چرخه روزانه ۱۲ ساعت روشنایی و ۱۲ ساعت تاریکی در دوران تکوین جنینی منجر به کاهش پایدار در میزان ترس و بهبود سلامت پا در بلندمدت شد. این اثرات پیامدهای اقتصادی قابلتوجهی دارند، از جمله تأثیر بر زمان عرضه طیور به بازار.
جدول ۲: اثرات نور بر رفاه و تولید
|
منبع |
نوع نور/شدت |
اثر بر ترس/رفتار |
اثر بر تولید |
|
Deep et al., ۲۰۲۳ |
سبز/قرمز (طول موج بلند) |
کاهش TI، نوک زدن به پر کمتر |
افزایش ۱۰% جوجهدرآوری |
|
Ncho et al., ۲۰۲۴ |
۲۰ لوکس (اولیه) |
فعالیت متوسط، رفاه بالا |
رشد بهینه، FCR پایین |
|
Wang et al., ۲۰۲۵ |
۲۳L:۱D (پیوسته) |
افزایش استرس، اختلال ریتم |
کاهش ۱۵% وزن، کیفیت گوشت پایین |
تعاملات عوامل استرسزا
استرسزاهای غیرزیستی اغلب بهصورت ترکیبی عمل میکنند و اثرات تشدیدکننده دارند. برای مثال، ترکیب استرس گرمایی و صوتی (مانند نویز تجهیزات در دماهای بالا) سطوح کورتیکوسترون را تا ۴۰% افزایش میدهد و میکروبیوتا را مختل میکند. مطالعهای در ۲۰۲۴ نشان داد که تعامل دما و نویز بر رفتار جوجهها تأثیر دوگانه دارد. همچنین، ارتعاش + نور نامنظم میتواند رشد جنینی را ۲۰% کاهش دهد. بنابراین جلوگیری از وقوع هرکدام از استرسها در طیور بصورت جداگانه، تأثیرات همافزایی آنها را نیز خواهد کاست.
استراتژیهای مدیریت و کاهش استرس
برای کاهش استرس، استراتژیهای زیر پیشنهاد میشود:
دما: استفاده از سیستمهای خنککننده تبخیری و مکملهای آنتیاکسیدان (مانند ویتامین E، ۲۰۰ IU/kg جیره) برای کاهش ROS تا ۳۰% و در استرس سرمایی، عایقبندی سالنها و جیرههای پرانرژی (افزایش ۱۰% کالری).
صدا و ارتعاش: عادتدهی تدریجی (desensitization) با صداهای کنترلشده از هفته اول؛ عایقبندی تجهیزات برای کاهش نویز به <۸۰ dB و ارتعاش <۵ Hz
نور: برنامههای نوری با ۱۶L:۸D برای بهبود ریتم شبانهروزی؛ شدت ۵-۲۰ لوکس برای جلوگیری از استرس. استفاده از طول موج سبز برای کاهش ترس تا ۲۵%.
اقتصادی و رفاه: این استراتژیها میتوانند ضرر ۲۰% تولید را به ۵% کاهش دهند، با ROI ۱۵۰% (نسبت سود و هزینه) در مزارع صنعتی (بر اساس مدلهای FAO ۲۰۲۵). نظارت با سنسورهای IoT برای پیشبینی استرس.
نتیجهگیری
اگرچه مقالات علمی موجود در زمینه استرس هنوز نامتوازن و نابرابر است، اما کمک چشمگیری به درک فیزیولوژی عمومی استرس کرده است. تنشهای غیرزیستی میتوانند موجب تغییر در متابولیسم، رفتار، و ایمنی گردند. استرس مزمن میتواند منجر به تغییرات نوروبیولوژیکی گردد که بر ساختار مغز تأثیرگذارند. در حالیکه عوامل بیماریزا (پاتوژنها) را میتوان با اقدامات پیشگیرانه رایج کنترل کرد، عوامل غیرزیستی مانند دما و نویز کنترلناپذیرتر و دشوارتر هستند.
پیشبینی میشود که تغییرات اقلیمی این مشکلات را وخیمتر کند، بهطوریکه در آینده نوسانات حرارتی شدیدتر (تا ۳۰% افزایش تا ۲۰۵۰) و اختلالات صوتی بیشتری رخ خواهد داد. علاوه بر این، اختلالات در برنامههای نوردهی یا سطوح ناکافی شدت نور میتوانند عملکرد تولیدی مرغهای تخمگذار را بهطور منفی تحت تأثیر قرار دهند. گرچه پژوهشها پیرامون تنشهای مرتبط با دما پیشرفت چشمگیری داشتهاند، اما درک دقیق و جزئی از تمامی تنشهای غیرزیستی برای دستیابی به سامانههای پایدار تولید طیور ضروری است. جامعه علمی باید تعاملات پیچیده میان طیور و محیط زیست آنها را نیز مورد بررسی قرار دهد تا بتواند راهکارهای مؤثر و کاربردی برای مدیریت استرسهای محیطی ارائه نماید.
گردآورنده: دکتر محمدرضا قهرمانیان
بازبینی و ویرایش: مهندس رضا یزدی
فهرست منابع
- Ncho, H. S., et al. (۲۰۲۴). *Abiotic stressors in poultry production: A comprehensive review*. Journal ofAnimal Physiology and Animal Nutrition, 108(3), 456-478.
- Wang, Y., et al. (۲۰۲۵). *Heat stress and the chicken gastrointestinal microbiota*. Journal of Animal Science and Biotechnology, 16(1), 12.
- Iraqi, M. M., et al. (۲۰۲۴). *Effects of abiotic stressors on poultry immunity*. Poultry Science, 103(2), 200-215.
- Zhou, Y., et al. (۲۰۲۱/بهروزرسانی ۲۰۲۵). *Cold stress impacts on broiler performance*. Animals, 11(5), 1345.
- FAO. (۲۰۲۵). *Climate change and poultry production: Economic models*. Food and Agriculture Organization Report.
- Deep, A., et al. (۲۰۲۳). *Lighting wavelengths and fear response in broilers*. British Poultry Science, 64(4), 389-397.






































































024532531.jpg?v=3Me6NG2_nIZ8WXVravM98a_fqZqLFWFs2xiJ9PUZq4A)


















































005501674.jpg?v=Tf8WTPxBKykjrdsOHwFqWCuAnE5XTw8TMCzymNi5ch0)


002226069.jpg?v=-9B5a0CcDYIVH1NSveV6FTmGe2r-kE3pbZhcGikxMKU)









005240285.jpg?v=pa7qIxfeulTU1ZxrGmmvOYjPl8H33JSqprB8FsfmDck)
003421766.jpg?v=Qkjs3wy10ezXHrlmr4Add3ksNJdNok5OS8LvvTUG8BU)
001716576.jpg?v=3QBhhkvX4x6nsPxUljY0c_2vOG_Qooasb9umgZ0Zy0M)